En las costas Venezolanas, así como en muchas otras áreas del Caribe y de zonas tropicales, es común encontrar octocorales acompañando arrecifes de corales pétreos (Figs. 1). Este es un hecho frecuente, que involucra a todas o casi todas las especies de estos organismos, así como a todos los tipos de formaciones coralinas. No obstante, en ciertas áreas, normalmente zonas expuestas a corrientes, nos encontramos formaciones exclusivas de octocorales o donde ellos representan el 80% o más de la cobertura de coral vivo (Fig. 2). Estos sistemas se desarrollan normalmente sobre plataformas de coral fósil o roca.

Figura 1. Fotografía de un octocoral en el borde de un arrecife, donde comienza el lecho arenoso.

Figura 2. Imágenes submarinas de sectores donde el sustrato está dominado por octocorales.
Para tratar de probar si esta dominancia de octocorales afecta significativamente la composición y estructura del ensamble de peces asociados, se estudiaron dos sistemas cercanos: uno rocoso coralino y otro exclusivo de octocorales.
INTRODUCCIÓNDiversos autores han demostrado que, a lo largo del planeta, las características del hábitat influyen de manera directa sobre la estructura de las comunidades ícticas que allí se desarrollan (Grigg, 1994; Caley y St John, 1996; Garpe y Oehman, 2003; Bouchon-Navarro et al., 2005). La gran mayoría de estos trabajos para el estudio de ensambles de peces, se han centrado en evaluaciones de la ictiofauna asociada a fondos blandos arenosos, areno-fangosos y de praderas de hierbas marinas; o a estructuras más complejas como arrecifes coralinos, rocosos o rocoso-coralinos (Sale, 1991, Larkum et al., 2006, Félix et al., 2007).
Escasas investigaciones tienen como foco de interés el análisis de la ictiofauna acompañante de sistemas de octocorales, a pesar de que se ha señalado la importancia de este hábitat como zona de refugio de peces (Risk et al.,1998). Los pocos trabajos en peces relacionados con fondos dominados por octocorales (gorgonios), son realizados principalmente en plataformas de aguas profundas, por debajo de los 50 m (Pirtle 2005). En espacios de menor profundidad, vale la pena mencionar la contribución de Syms y Jones (2001), donde se muestra el efecto que se causa sobre la comunidad de peces al eliminar los corales blandos en un sistema de arrecifes. El resto de las investigaciones, se restringen a casos de especies particulares, por ejemplo la depredación de Chaetodon capistratus sobre octocorales gorgónidos en el Caribe (Lasker, 1985).
En tal sentido, y viendo además que en Venezuela no existían registros sobre peces asociados a octocorales, se decidió evaluar dos sistemas ecológicos cercanos, uno dominado por corales blandos y otro rocoso coralino, bajo la hipótesis de que las diferencias estructurales entre ambos sustratos deberían marcar cambios importantes en la ictiofauna que en ellos habita.
METODOLOGÍA
En el Bajo Las Caracas, un promontorio submarino del estado Sucre, Venezuela, se escogieron dos sustratos adyacentes pero disímiles en su estructura para la realización del estudio. El primero es un promontorio de arrecife constituido por rocas y corales pétreos, con profundidades que oscilan entre los 3 m y los 6 m, presentando una hilera longitudinal de rocas altas (2 a 3 m), con numerosas oquedades, generando una importante complejidad estructural al espacio. Paralelo a este arrecife (a unos 5 m de distancia) se localiza el sustrato de octocorales, con una profundidad media de unos 6 m, caracterizado por la ausencia de corales pétreos, con corales blandos sobre una base dura probablemente de coral fósil o de roca (Fig. 3).

Figura 3. Imágenes submarinas de los dos sistemas evaluados: Octocorales (arriba) y arrecife rocoso coralino (abajo).
Se efectuaron censos visuales mensuales, durante un año, en dos transectos, uno en cada arrecife. Se utilizó una modificación del protocolo Atlantic and Gulf Rapid Reef Assessment (AGRRA) (Ginsburg 2000), que incluye el conteo de todas las especies observadas. Igualmente, la longitud de los transectos fue ampliada a 40 x 2 m, con relación a la establecida por AGRRA, para comprender un área más representativa del arrecife. La identificación de las especies se realizó utilizando las descripciones de Humann y Deloach (2002). Para establecer la similaridad entre los transectos, respecto al número de especies, se efectuó el cálculo de disimilaridad de Bray Curtis (1957), utilizando el programa MVSP (Servicio de Computación Kovac, 2006).
Debido a que los datos no presentaban normalidad en su distribución, se procedió a realizar un análisis no paramétrico de Kruskall-Wallis, mediante el programa Statgraphics Plus 4.1, para comparar los promedios anuales de la diversidad, la equitatividad, la riqueza y la abundancia de las especies de peces entre los dos arrecifes. Para visualizar la variación de las abundancias de las especies a través del año y entre los arrecifes, se aplicó un análisis de componentes principales (ACP) para cada ecosistema, mediante el programa MVSP, considerando los meses como los casos y a las especies como variables, descartando las pelágicas así como aquellas cuyas abundancias representaron menos del 10% de la abundancia total.
RESULTADOS Y DISCUSIÓN
En los dos arrecifes se censaron 5.655 individuos, pertenecientes a 84 especies de peces, incluidas en 32 familias; de las cuales, 80 fueron observadas asociadas al transecto rocoso coralino y 41 especies en el transecto de los octocorales. Todos los peces observados fueron óseos (Osteichthyes), con excepción del tiburón gata Ginglymostoma cirratum: Chondrichthyes, Rhincodontidae), visto en el arrecife pétreo.
Las familias con mayor importancia numérica en el arrecife pétreo fueron Scaridae (15,22%), Haemulidae (14,18%), Carangidae (13,88%) y Labridae (12,59%); mientras que en los octocorales, las familias con mayor abundancia fueron Clupeidae (31,69%) Carangidae (26,56%), Scaridae (17,33%), Haemulidae (9,13%) y Aulostomidae (5,78%) (Fig. 4).

Figura 4. Importancia relativa de las principales familias observadas en dos arrecifes del Bajo Las Caracas, entre julio de 2005 y junio de 2006.
Las especies más abundantes observadas en el arrecife pétreo fueron Decapterus punctatus (13,06%), Thalassoma bifasciatum (11,64%), Haemulon chrysargyreum (10,98%) y Acanthemblemaria sp. (7,52%); mientras que en los octocorales, las especies con mayor importancia numérica relativa fueron: Sardinella aurita (31,69%) Decapterus punctatus (25,07%), Sparisoma aurofrenatum (10,16%) y Aulostomus maculatus (5,78%) (Fig. 5).

Figura 5. Importancia relativa de las principales especies observadas en dos arrecifes del Bajo Las Caracas, entre julio de 2005 y junio de 2006.
Destaca la presencia de Aulostomidae y de su especie representativa, Aulostomus maculatus, dentro del grupo con mayor importancia numérica relativa sobre los octocorales. A. maculatus es registrada frecuentemente en arrecifes pétreos pero en densidades bajas; mientras que en los octocorales ocupa el tercer puesto en importancia numérica. Este pez asume coloración críptica en posición vertical entre las ramas de corales blandos, por lo que dicho ambiente ofrece un refugio ideal, así como un lugar de acecho apropiado para sus actividades predatorias, un hecho ya señalado por Ramírez y Cervigón (2003).
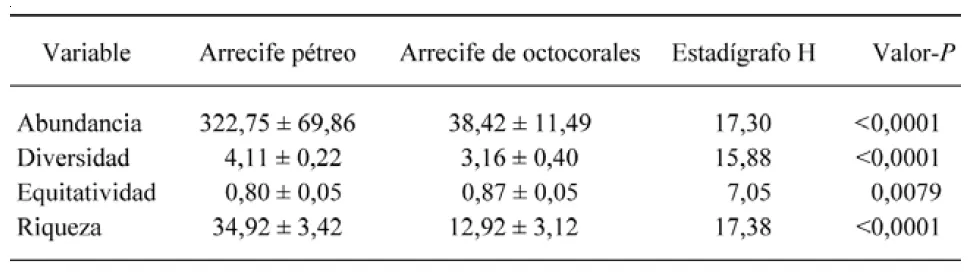
El análisis de Disimilaridad de Bray Curtis arrojó un valor de distancia de 0,758 entre los dos arrecifes, por lo que se consideran significativamente distintos respecto a las abundancias de las especies de peces. El análisis de Kruskal-Wallis arrojó diferencias significativas respecto a la abundancia, la diversidad, la equitatividad y la riqueza de especies entre los dos arrecifes (Tabla I).
Tabla I. Resultados de las pruebas de Kruskal-Wallis para evaluar las diferencias entre los dos arrecifes estudiados respecto a componentes comunitarios.
Los resultados del análisis de componentes principales indican una mayor dispersión de la abundancia mensual de las especies en el arrecife de octocorales respecto del pétreo (Figura 6). Tal situación pudiera estar reflejando que el ensamble de peces en los corales blandos es menos estable, con especies que fluctúan anualmente entre este sustrato y zonas foráneas. Esta hipótesis se refuerza cuando se observa la presencia de los taxa durante los meses de muestreo, ya que en los octocorales, la mayoría de las especies hacen un uso accidental del espacio (53,66%) o son accesorias (21,95%); y sólo una cuarta parte se presenta de manera constante (24,39%) (Figura 7). De las 41 especies presentes en este sustrato, sólo Aulostomus maculatus, Sparisoma aurofrenatum y Sparisoma viride; estuvieron en todos los meses de muestreo.
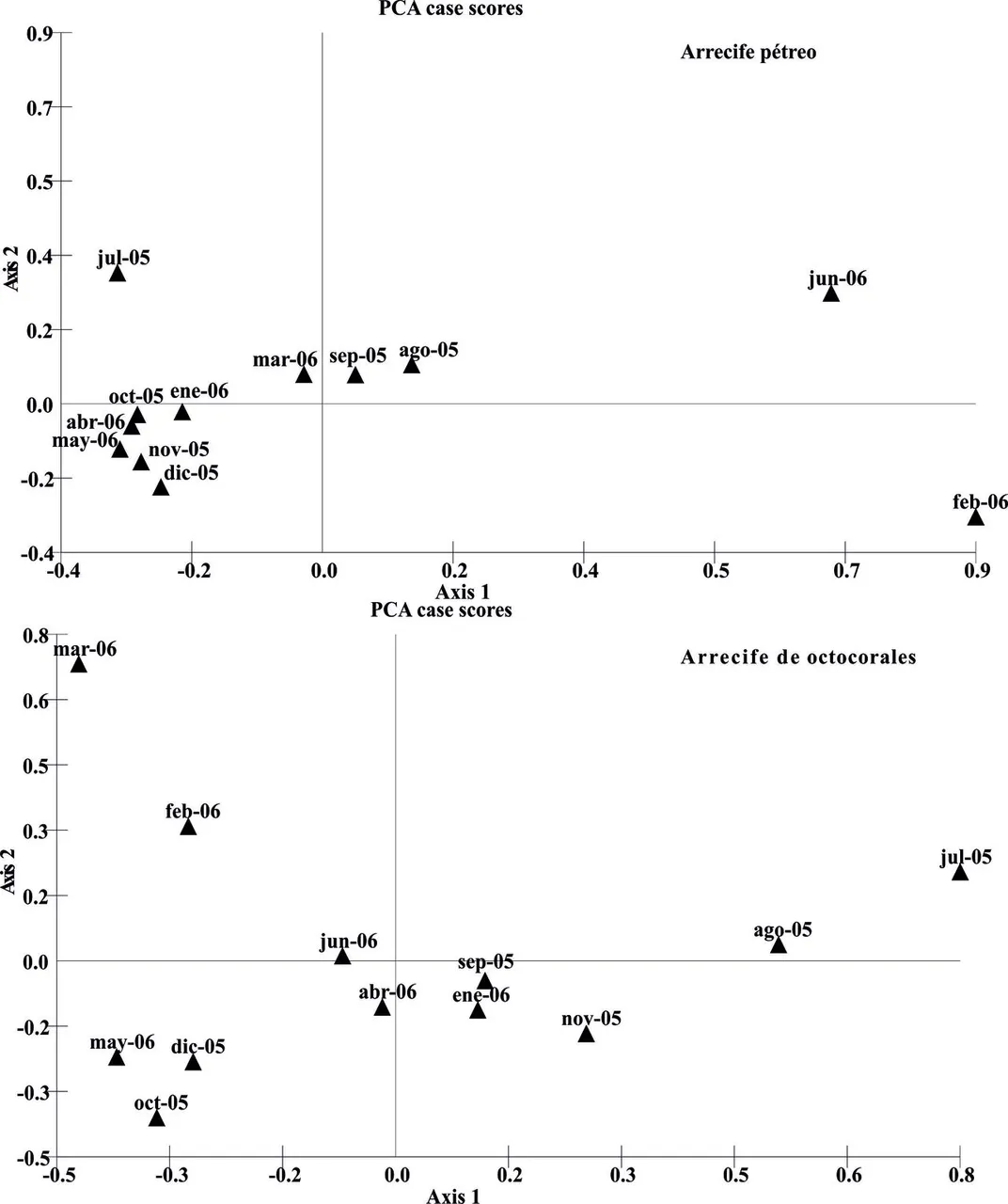
Figura 6. Scatter plot del Análisis de componentes principales para relacionar los meses de muestreo con las abundancias de las especies en dos arrecifes del Bajo Las Caracas, entre julio de 2005 y junio de 2006.
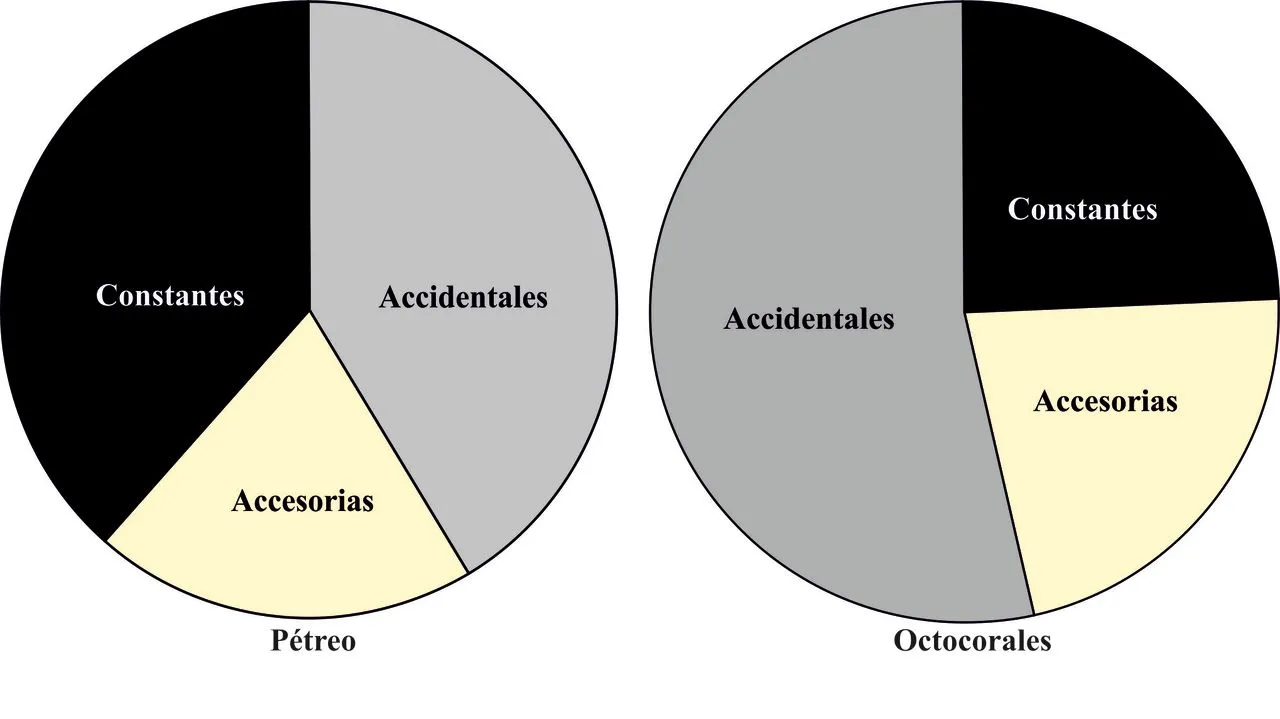
Figura 7. Representación gráfica de los porcentajes para los valores de constancia de las especies observadas en dos arrecifes del Bajo Las Caracas, entre julio de 2005 y junio de 2006.
Resalta además que especies como Acanthurus coeruleus, Haemulon flavolineatum, Microspathodon chrysurus, Odontoscion dentex, Scarus vetula, Stegastes partitus, Mulloidichthys martinicus, Haemulon carbonarium y Cephalopholis cruentata, que estuvieron de manera constante sobre el arrecife pétreo a lo largo de todo el año de muestreo, no fueron observadas ni siquiera accidentalmente sobre los octocorales.
Al analizar estos dos sistemas de sustratos, adyacentes pero estructuralmente distintos, se observa que el arrecife pétreo es más diverso y con mayor riqueza que el sector de octocorales. Esto pudiera estar determinado por la alta heterogeneidad estructural que ofrecen las grandes rocas con sus innumerables oquedades, generando una disponibilidad de refugio para los peces más elevada (Friedlander y Parrish, 1998; Syms y Jones, 2000) respecto al también heterogéneo, pero menos complejo, sistema de ramificaciones en los octocorales. Además, sobre los octocorales son limitados los epibiontes que se forman, debido a sus defensas químicas así como su superficie móvil y menos extensa. Estas características, en conjunto, traen como consecuencia una menor cantidad de alimento disponible y, por ende, un número más bajo de especies de peces asociadas. Por lo tanto, los recursos en los octocorales se encuentran más limitados, por lo que las presiones selectivas deberían ejercer una mayor fuerza competitiva, lo cual pudiera influir en una ocupación selectiva del espacio por especies apropiadamente adaptadas.
Todavía faltan estudios en áreas similares, pero con base en las evidencias ícticas y entendiendo la necesidad de ampliar a otras zonas y otros taxa asociados, se propone considerar a los arrecifes de corales blandos como un tipo particular de ecosistema que, conjuntamente con las praderas de fanerógamas marinas, las raíces sumergidas de mangle, los arrecifes de coral tradicionales, los arrecifes rocosos y los diferentes tipos de fondos blandos, entre otros, estarían representando la diversidad de ecosistemas que conforman las aguas sumergidas venezolanas.
REFERENCIAS
Bouchon-Navarro Y, C Bouchon, M Louis & P Legendre. 2005. Biogeographic patterns of coastal fish assemblages in the West Indies. Journal of Experimental Marine Biology and Ecology, 315: 31-47.
Bray, J. R. Y J. T. Curtis. 1957. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 27: 325–349.
Caley MJ y J St John. 1996. Refuge availability structures assemblages of tropical reef fishes. Journal of Animal Ecology, 65: 414-428.
Fariña A., E. Méndez, S. Sant y E. Zapata 2008. Diferencias en la composición de especies de peces entre un arrecife rocoso-coralino y uno de octocorales en el Bajo Las Caracas, estado Sucre, Venezuela. Bol. Centro Invest. Biol., 42(2): 165–185.
Fariña A. y E. Méndez. 2009. Variación estacional de la estructura comunitaria de peces en dos arrecifes: rocoso-coralino y de octocorales, en el Bajo Las Caracas, Venezuela. Rev. Biol. Mar. Oceanog., 44(1): 153-162.
Félix, F. C., H. L. Spach, P. S. Moro, C. W. Hackradt, G. M. Nogueira De Queiroz Y M. Hostim-Silva. 2007. Ichthyofauna composition across a wave-energy gradient on southern Brazil beaches. Brazilian J. Oceanography 55(4): 281–292.
Friedlander, A. M. y Parrish, J. D. 1998. Habitat characteristics affecting fish assemblages on a Hawaiian coral reef. J. Exp. Mar. Biol. 224: 1-30.
Garpe KC & MC Oehman. 2003. Coral and fish distribution patterns in Mafia Island Marine Park, Tanzania: fish-habitat interactions. Hydrobiologia 498(1-3): 191-211.
Ginsburg, R. 2000. “AGRRA protocol. Atlantic and Gulf Rapid Reef Assessment”. Universidad de Miami, RSMAS/AGRRA, 4600 Rickenbacker Causeway, Miami FL 33149, USA. Disponible en http://coral.aoml.noaa.gov/agra. Visitado el 08 de marzo de 2007.
Grigg R. 1994. Effects of sewage discharge, fishing pressure and habitat complexity on coral ecosystems and reef fishes in Hawaii. Marine Ecology Progress Series, 103: 25-34.
Humann P & N Deloach. 2002. Reef Fish Identification, 512 pp. New World Publications, Jacksonville.
Larkum, A. W. D., R. J. Orth Y C. M. Duarte. 2006. Seagrasses: Biology, Ecology and Conservation. Springer, Netherlands. 691 pp.
Lasker, H. R. 1985. Prey preferences and browsing pressure of the butterflyfish Chaetodon capistratus on Caribbean gorgonians. Mar. Ecol. Prog. Ser. 21: 213–220.
Pirtle J. L. 2005. Habitat-based assessment of structure-forming megafaunal invertebrates and fishes on Cordell Bank, California. Tesis de Maestría, Universidad del Estado de Washington, Vancouver,, USA, 74 pp.
Ramírez, H. y Cervigón, F. 2003. Peces del archipiélago Los Roques. Caracas: Agencia Española de Cooperación Internacional, Araucaria.
Risk, M. J., D. E. Mcallister Y L. Behnken. 1998. Conservation of cold-and warm-water seafans: Threatened ancient gorgonian groves. Sea Wind 10(4): 20–22.
Sale, P. F. 1991. The ecology of fishes on coral reefs. Academic Press, San Diego, California, USA, 754 pp.
Servicio De Computación KOVAC. 2006. Multi Variate Statistical Package (MVSP), Versión 3.13n. Kovach Computing Services: The easy and inexpensive way to analyze your data. Disponible en http://www.kovcomp.com. Visitado el 02 de enero de 2007.
Syms, C. Y G. P. Jones. 2001. Soft corals exert no direct effects on coral reef fish assemblages. Oecologia 127: 560–571.</div>
SI ERES CIENTÍFICO O TE GUSTA LA CIENCIA, TE INVITO A UNIRTE A NUESTRO CANAL EN DISCORD SIGUIENDO ESTE ENLACE: https://discord.gg/6fZgub Y A NUESTRA COMUNIDAD @STEM-ESPANOL Y @STEEMSTEM
